



Az információfeldolgozás titkai és megértésének nehézségei
Az eLife folyóiratban, Magó Ádám és Kis Noémi elsőszerzőségével jelent meg Makara Judit csoportjának cikke. A technikailag is kihívást jelentő projekt feltárta a memóriafolyamatokban érdekelt, eltérő morfológiájú piramissejtek néhány titkát.
Az eLife folyóiratban, Magó Ádám és Kis Noémi elsőszerzőségével jelent meg Makara Judit csoportjának cikke. A technikailag is kihívást jelentő, "nyomozós" projektnek sikerült feltárnia a memóriafolyamatokban érdekelt, eltérő morfológiájú piramissejtek néhány titkát.
Egy általános meghatározás szerint a tudományos ismeretterjesztés a társadalom egészének érdekében végzett, közoktatási rendszeren kívüli oktatás. Bár sok kiváló kutató is részt vesz benne, ez nem kutatási tevékenység és nem tudomány. Azt ugyan jó észben tartani, hogy Isaac Newton 1687-ben publikált, egész Európában páratlan sikert aratott, saját korában is számos kiadást megélt, "A természetfilozófia matematikai alapjai" (Philosophiae Naturalis Principia Mathematica) című könyve is ismeretterjesztési célt szolgált, de ez a színvonal semmiképp nem várható el egy mai ismeretterjesztőtől.
Newton latinul írta könyvét, hogy minél többen olvassák, és az egész, magát műveltnek tartó világ megértse. Ma, amikor a tudomány nemzetközi nyelve, a lingua academica az angol, és nem csak nálunk, sajnos másutt sem általános, hogy igény lenne az anyanyelv, és azon belül a tudományos szaknyelv ápolására, fejlesztésére is, egyre nehezebb fontos eredmények érthető közvetítése a tudomány iránt érdeklődő, de azt nem művelők számára.
Makara Judit, az eLife folyóiratban cikk vezető kutatója azonban igyekezett mindent megtenni azért, hogy minél többen megértsék a Magó Ádámmal, Kis Noémivel és Lükő Balázzsal együtt végzett munkájuk célját és eredményeit.
De kövessük a maga korában sokak által olvasott Lord Byron módszerét - My way is to begin with the beginning - és kezdjük az elején!
Makara Judit:
- Ez a cikk egy hosszabb kutatási programhoz tartozik, amelyben az agy hippokampusz nevű részének CA3 régiójában található piramissejteket vizsgáljuk azzal a céllal, hogy megértsük, hogy a sejtek elektromos és jelintegrációs tulajdonságai hogyan illeszkednek ennek a régiónak az információfeldolgozásban játszott szerepéhez.
- Az információfeldolgozás korunk egyik "varázsszava". Nagyon rövidre fogva, mit jelent ez a hippokampusz CA3 régiója esetében?
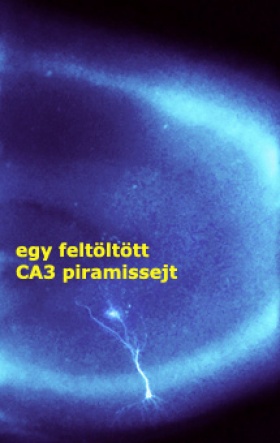
- Részben az, hogy a terület kísérleti vizsgálhatósága nem túl kedvező, mivel a sejtek felnőtt rágcsálókból készített agyszeletekben sérülékenyek, hamar elpusztulnak. Jóval több ismeretünk van a CA1 piramissejtek dendritjeiről, ezek a sejtek ugyanis jól fenntarthatók felnőtt állatból készített agyszeletben is, dendritfiziológiai mérések is könnyen végezhetők rajtuk. Így már kb. 10-15 éve is viszonylag sokat tudtunk a dendritikus integrációról a CA1 területén, és sokan feltételezték, hogy ezek a tulajdonságok átemelhetőek lesznek a többi piramissejt típusra is.
- A piramissejt név ezeknek a sejteknek az alakjára, azaz morfológiájára utal. Milyen különbség van köztük?
- A két terület piramissejtjeinek dendritfája elég látványosan különböző.
A CA1 piramissejtnek tipikusan egy (vagy egyszer elágazódó) hosszú fő apikális dendritága van, amely jól hozzáférhető az elektrofiziológiai mérések, az úgynevezett patch clamp vizsgálatok számára. A CA3 piramissejtek fő dendritjei hamarabb elágaznak több vékonyabb ágra, amelyek nehezebben megközelíthetők. Részben ez a technikai nehézség az oka annak, hogy a sejttesttől távoli CA3 dendriteken közvetlen patch clamp kísérleteket rajtunk kívül kevesen végeztek. A mi nemrégi eredményeink azonban arra utaltak, hogy fontos lenne megismerni a dendritikus Ca2+ spike-ok tulajdonságait ezekben a sejtekben.
- Miért fordult érdeklődésetek a Ca2+ spike-ok felé?
- Előző kísérleteinkben a CA3 piramissejtek tüzelési tulajdonságait a sejttestben mérve vizsgáltuk, és azt láttuk, hogy a sejtek áraminjekcióra adott tüzelési válaszai nagyon heterogének. Egyes sejtek viszonylag szabályos időközönként tüzeltek egyedi akciós potenciálokat, míg másokra jellemző volt, hogy inkább jellegzetes sorozattüzeléseket, ún. komplex-spike burst-öket (kiejtés: bɜːst) produkáltak. Ez esetben egy gyors akcióspotenciál-sorozat egy lassú depolarizációs feszültséghullámra ül rá, amelyért jellemzően a dendritekben kialakuló Ca2+ spike-okat tartják felelősnek.
Kíváncsiak voltunk, vajon a dendritikus tulajdonságok megmagyarázzák-e azt, miért ilyen heterogén ezekben a sejtekben a burst tüzelési hajlandóság, és közvetlenül a dendritekből kezdtük el mérni, hogyan reagálnak depolarizációt kiváltó áraminjekciókra. Azt feltételeztük, hogy a burst-ölő sejtek dendritjeiben könnyen kiválthatóak a (tipikusan lassú) Ca2+ spike-ok, de a nem burstölő sejtekben esetleg nincsenek meg, vagy csak nehezen aktiválhatók a szükséges Ca2+ spike mechanizmusok.
- Ez valóban logikusnak tűnik. . .
- Meglepetésünkre, mégsem ezt találtuk: az áraminjekcióval kiváltott dendritikus regeneratív válaszok e helyett zavarba ejtően sokfélék voltak. Különféle regeneratív válaszformákat láttunk: gyorsat, lassút, minden dendritben kicsit mást, ráadásul az eredmény attól is függött, milyen messze mértünk a sejttesttől, és milyen erősségű volt a depolarizáció. Ne feledjük, hogy a dendritekben nem csak Ca2+ spike-okra, hanem lokális Na+ spike-okra és a sejttestről visszaterjedő akciós potenciálokra is számítanunk kellett.
- Ezt olyan lelkesen meséled, mintha kifejezetten élveztétek volna, hogy a helyzet ahelyett, hogy egyszerűbb és érthetőbb lett volna, egyre átláthatatlanabb lett. Legalábbis egy hozzá nem értő számára. . .
- Valóban nem volt egyszerű. Ez egy nagyon érdekes, „nyomozós” projekt lett, sok fejtörésbe került, amíg a mozaik elemei a helyükre kerültek.
Először is ahhoz, hogy megértsük, mi honnan jön, mi mit eredményez, kénytelenek voltunk olyan, meglehetősen nehéz kísérleteket is végezni, amikor a sejttesten és a dendriten egyszerre mérünk egy-egy elektródával. Ebben az esetben ugyanis a jelek nagysága, kinetikája és a kiváltott dendritikus Ca2+ válaszok alapján könnyebben azonosítani tudjuk a különböző regeneratív kisülések eredetét.
A sok kísérlet eredményeként végül a dendritikus Ca2+ spike-okat két fő csoportba tudtuk sorolni. Amellett, hogy találtunk viszonylag lassú Ca2+ spike-okat, melyeket a dendritekbe visszaterjedő akciós potenciálok váltottak ki és gyakran tipikus burst-öket eredményeztek, felfedeztünk egy új Ca2+ spike típust, ami lokálisan a dendritben indult, kiváltó szomatikus aktivitás nélkül. Ez a Ca2+ spike fajta szokatlanul gyors (néhány miliszekundum) lefutású volt, és ha szomatikus választ váltott ki, az mindig kizárólag csak egy akciós potenciál volt, nem pedig burst.

- Tehát sikerült kinyomozni a "magányos" akciós potenciálok eredetét?
- Részben igen. Kiderült, hogy a szabályosan tüzelő sejtek dendritjei is lehetnek nagyon is aktívak, csak éppen egy másik típusú, gyors Ca2+ spike-kal (is) rendelkeznek, amelyek nem a megszokott, "burstölő" viselkedést váltják ki a szómán (sejttesten). Ez ellene megy annak, amit a Ca2+ kisülésekről eddig gondoltunk, mert eddig az volt az általános elképzelés, hogy a Ca2+ spike-ok burst-szerű tüzelést eredményeznek, az új típusú gyors spike-ok azonban önmagukban inkább a burst kialakulása ellen hatnak.
- Adott tehát kétféle tüzelési mintázat. Mi a jellemző rájuk?
- A burstnek nevezett sorozatkisülést megbízhatóbb információátadási formának tekinthetjük, mivel a beidegzett piramissejtek általában erősebben válaszolnak sorozatszerű szinaptikus bemenetre. Ezzel szemben a gyors Ca2+ spike által kiváltott akciós potenciál a szómán ugyanolyannak látszik mint egy normál, megszokott egyedi akciós potenciál.
- Munkátok egyik értéke az is, hogy bár kísérleteiteket patkány idegsejteken végeztétek, találtatok kapcsolódási pontot az emberi idegsejtekkel is.
- Ez valóban érdekes aspektusa volt a kutatásnak.
Két éve, amikor már nagyjából láttuk, milyen különböző Ca2+ spike-ok fordulhatnak elő a CA3 piramissejt dendritekben, megjelent egy cikk a Science-ben Matthew Larkum csoportjától, ahol műtéti szövetből származó, emberi kortikális 2/3-as rétegi piramissejt dendritekből vezettek el, és nagyon hasonló gyors Ca2+ spike-okat láttak ők is. A cikk nagy érdeklődést keltett, és sokan arra következtettek, hogy íme, valami, ami talán csak emberben található. Mi azonban akkor már tudtuk, hogy hasonló spike-ok nem csak az emberi, de a patkány idegsejtekben is előfordulhatnak, és nem csak a neocortexben, hanem a hippokampuszban.
- Nem fajok, mint először gondolták, hanem inkább sejttípusok között van különbség. De mindennek oka van: mire jó ez?
- Ma nem tudjuk még, de nagyon szeretnénk megérteni, mi lehet a komputációs szerepe ezeknek a spike-oknak a patkány agyban, hiszen a hasonlóságot kihasználva esetleg extrapolálni lehetne az emberi agyban betöltött szerepre is.
- Most merre indultok tovább?
- Az egyik fő irányt a biofizikai mechanizmusok tisztázása jelenti, hiszen a felfedezésünk szükségszerűen felveti a kérdést, hogy ioncsatorna okozta különbségek okozzák-e a különböző Ca2+ spike típusokat, és hogy ezek mennyire-hogyan függenek a morfológiától.
Az egész munkához konceptuális hátteret ad a CA3 terület lehetséges szerepe a memória folyamatokban. A cikkben megjelent munkánk természetesen nem a memóriáról szól, hiszen annál sokkal speciálisabb, sejt szintű vizsgálatokat végeztünk. Úgy gondolom, fontos eredmény lenne, ha a különböző spike típusok tanulásban és memóriában betöltött szerepét megérthetnénk. Ehhez élő, memóriafeladatot végző állatok agyában is próbáljuk azonosítani a szeletben megismert folyamatokat.
Ujfalussy Balázs csoportjával is tervezünk együttműködni abban, hogy a régió modellezésével jobban megértsük a különböző Ca2+ spike típusok szerepét, mikor-miben segíthet ez a heterogenitás és a különleges tulajdonságú dendritikus spike-ok.
- Már szóba került, hogy a kísérletek elvégzése a CA3 piramissejtek morfológiája miatt is nagy kihívást jelentett. Kiknek sikerült ezzel megbirkózniuk?
- Amikor megkezdtük a CA3 régió vizsgálatát, felkészültünk a technikai kihívásokra a dendritek nehéz megközelíthetősége és a sejtek sérülékenysége miatt is. Való igaz, technikailag nem könnyű, "rá kell érezni" a vékony dendritek patch-elésére, de ahhoz képest, amire számítottunk, elég jól tudtunk haladni. Ez elsősorban Magó Ádám érdeme, aki a mérések beállítását és a dendritkísérletek nagy részét végezte. Miután Ádám posztdoktorként külföldre ment, a munkát Kis Noémi PhD hallgató (a cikk megosztott első szerzője) folytatta, aki korábban már hozzájárult a Ca2+ spike-ok sejttestről történő vizsgálatához, majd megtanulta a nehéz dendritelvezetések végzését is. Lükő Balázs azokat az analízis modulokat fejlesztette ki, amelyekkel a spike-ok tulajdonságait elemeztük.