



Sokat kevésről vagy keveset sokról: az elektrofiziológiai módszerek palettája II.
A hálózatok duruzsolásának kihallgatására alkalmas módszerek után, ma az egyedi idegsejt viselkedését vizsgáló elektrofiziológiai módszerek kerülnek sorra.
A következő lépés az úgynevezett juxtacelluláris elvezetés. Juxta = közeli, azaz a sejtek közvetlen közeléből vezetünk el egy vékony üvegkapillárissal (~0.5µm).
Itt már csak egyetlen sejt aktivitását tudjuk vizsgálni és ha ügyesek vagyunk a vizsgálat végén a sejtet egy jelölőanyaggal a kapillárison keresztül fel is tudjuk tölteni, hogy később megvizsgálhassuk szerkezetét, típusát, összeköttetéseit.
Ha vékony üvegelektródánkat a sejt belsejébe szúrjuk minőségi ugrást hajtunk végre. Ilyenkor már sejtünkön kívül senkit nem hallunk, csak sejtünket, de azt jól. Közvetlenül tudjuk mérni a sejt membránpotenciálját, mely a nyugalmi potenciál -65mV-járól a gátlás -75mV-jára vagy az akciós potenciál csúcsának +15 mV-jára ugrik. Képesek vagyunk a sejtre érkező serkentő és gátló áramokat, valamint a sejt kimenetét, akciós potenciál sorozatait is pontosan mérni. Ezt az elvezetést hegyes elektródás intracelluláris (sejten belüli) elvezetésnek nevezik. A sejtek jelölése és azonosítása itt is elvégezhető. Elektrofiziológiai módszereink sorában ez az a lépés amikor a lelátón beszélgetésbe elegyedünk valakivel. Csak róla tudunk meg dolgokat, de sokat.
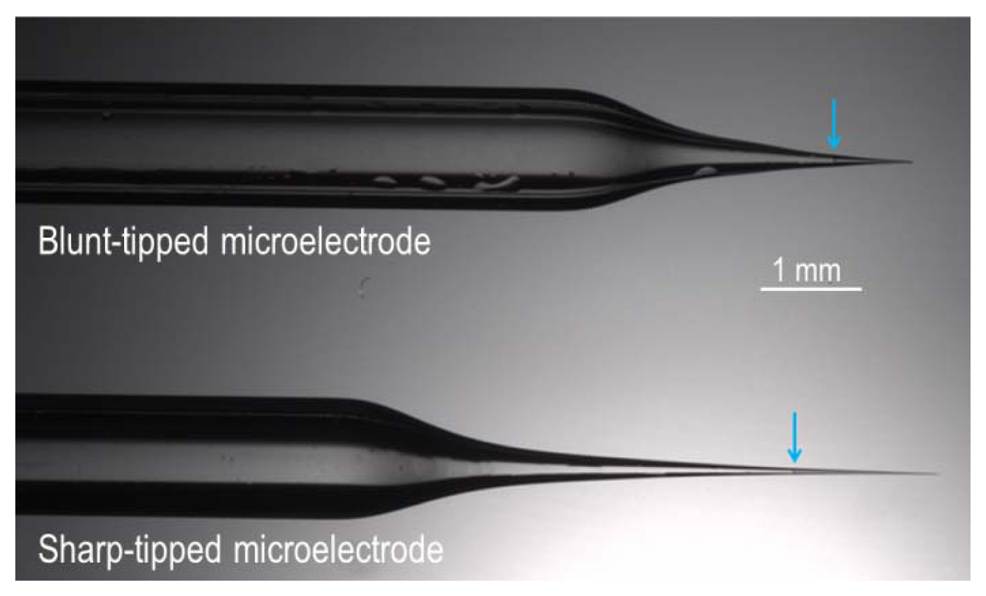
De ezzel még nem merült ki az elektrofiziológia repertoárja. Ha egy kicsit nagyobb szájátmérőjű (2-3 µm) elektródát használunk, akkor nem lukat szúrunk a sejtbe, hanem rátapadunk és egy jól záró kapcsolaton keresztül hozzáférünk a sejt belsejéhez. Egy ilyen elektródát használva nem csak a sejt membránpotenciálját mérhetjük, hanem ravasz erősítő rendszereket használva a sejtre érkező serkentő és gátló áramokat is elválaszthatjuk egymástól, illetve a sejtek jelintegrációjának szabályait is vizsgálhatjuk. Ezt az elvezetés típust hívják patch-clamp (folt-csapda) elvezetésnek és az áramok mérésére alkalmas módszert voltage-clamp (feszültség csapda) módszernek.
De még innen is van tovább. Ha elektródánkkal a sejt egy általunk kiválasztott részéhez tapadunk, majd azt leszakítjuk róla akkor az elektródához tapadt sejthártya darabban található ioncsatorna molekulákon keresztül folyó áramok tulajdonságait tanulmányozhatjuk. Így feltérképezhető, hogy az idegsejtek különböző részein milyen tulajdonságokkal rendelkező feszültség vagy átvivőanyag érzékeny molekulák találhatók. Ez a sokat kevésről vége a spektrumnak. Itt az egyetlen molekulán átfolyó áramok tulajdonságait vizsgálhatjuk és ezáltal a molekula működéséről vonhatunk le következtetéseket. Illetve felderíthetjük, hogy a sejtek felszínének eltérő részein (dendrit, sejttest, axon) milyen, a jelterjedésben és jelösszegződésben fontos molekulákból mennyi és milyen tulajdonságokkal található.
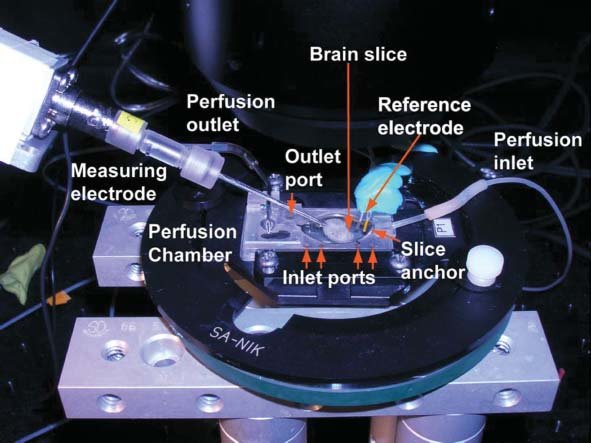
A mikroelektródás elvezetéseket gyakran nem élő, szabadon mozgó állatokból végzik, hiszen igen nehéz a mikrométeres sejtet és az elektródát egymáshoz képest rögzítve tartani egy mozgó kísérleti állat esetében. Megoldásként kidolgozták az In vitro (üvegben) elektrofiziológia módzsereit, ahol a vizsgált agyterüleből készített szeleteket tartanak életben megfelelő oldatokkal és ezek sejtjeiből vezetnek el egy vagy több mikroelektródával (bővebben lásd később).
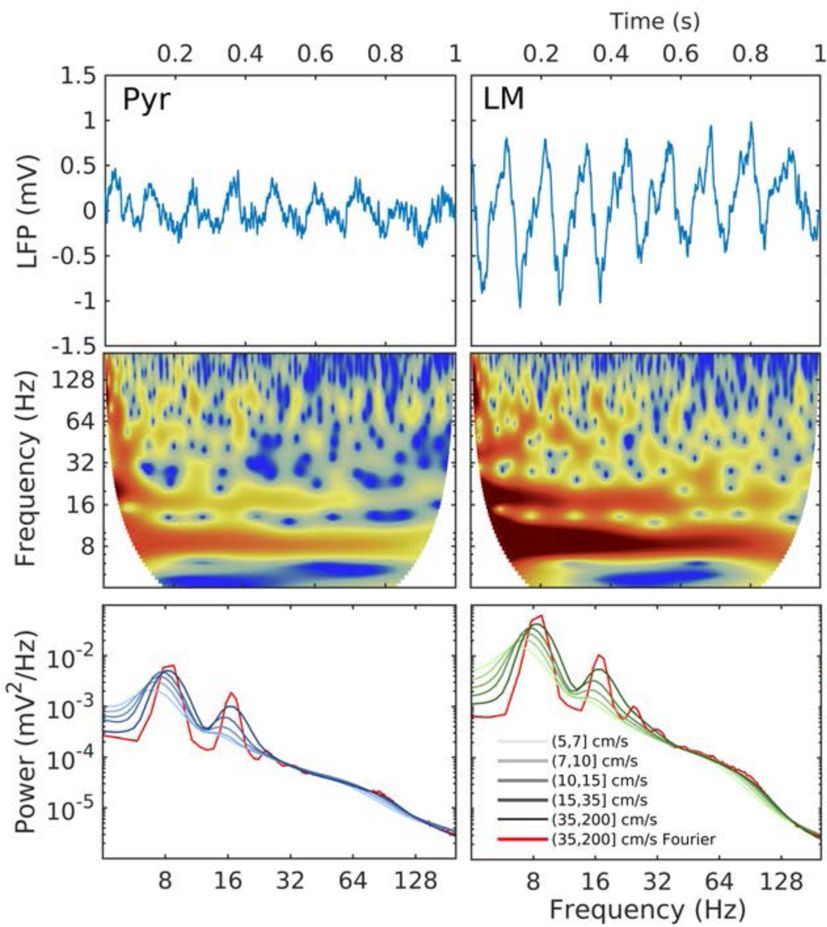
A módszereknek azonban csak egyik része az, hogy milyen elektródákat használunk, a másik az, hogy a jeleket milyen módszerekkel dolgozzuk fel. Az EEG jeleket frekvenciájuk szerint szétválogathatjuk. Ha több csatornánk van megnézhetjük, hogy ezek térben és időben hogyan függenek össze. Ha egy gyors feladatot adunk a kísérleti alanynak vagy elektromosan ingereljük az agyat megnézhetjük milyen időzítéssel és milyen lépcsőkben terjed az ingerület, mik a feldolgozás lépései. Ha nagyon sok csatornánk van, matematikusok és fizikusok által fejlesztett bonyolult statisztikai és jelfeldolgozási módszereket használunk. Nézhetünk összefüggéseket (korrelációk), nézhetjük a jelek mennyire egyszerre változnak (koherencia), de megnézhetjük a közös vagy eltérő információtartalmat és ezáltal a feltételezett kölcsönhatások irányát is becsülhetjük. A módszerek között ma már ismerősként köszöntik a gépi tanulást és a mesterséges neuronhálózatokat.
Egy másik módszert használva, a Mikroelektróda soros elvezetések esetében, ha az egymás melletti csatornák feszültségváltozásának változását kiszámítjuk megkaphatjuk azt, hogy mely agyi rétegekbe érkeznek serkentő vagy gátló áramok és ezeknek mi az időzítése. Ha a jel alacsony és magas frekvenciájú alkotóelemeinek összefüggését vizsgáljuk becsülhetjük, hogy az idegsejtek a rájuk érkező szinaptikus áramokat hogyan alakítják akciós potenciálokká, hogyan dolgozzák fel az információt.
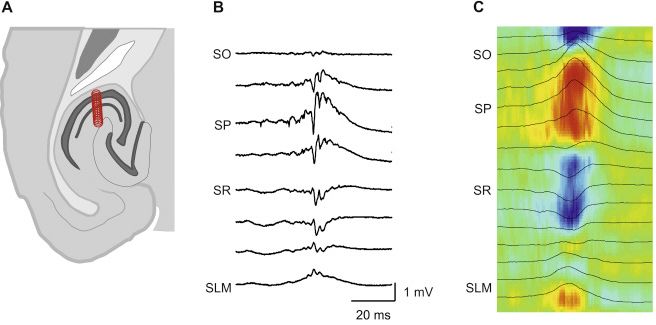
Az eltérő típusú módszereket kombinálhatjuk is. Ha a hálózat átlag viselkedését vizsgáló EEG-mezőpotenciál módszereket egyedi sejtek viselkedését megfigyelő módszerekkel (intracelluláris, patch clamp, multiunit) módszerekkel kombináljuk arról vonhatunk le következtetéseket, hogy az egyes azonosított sejtek hogyan járulnak hozzá a hálózat működésének alakításához. Azonosított sejtek esetében megtudhatjuk az egyes sejtek a csoport többi tagjához képest mikor működnek.
A továbbiakban részletesen beszélünk majd ezekről az elvezetési és elemző módszerekről. Addig is, ha bármi az eszetekbe jut kérdezzetek a szurkeallomany@koki.hu email címen!